La sintesi degli acidi grassi, nota anche come lipogenesi o liposintesi, rappresenta una via metabolica cellulare fondamentale. Questo processo parte da molecole di acetil-CoA per generare acido palmitico, un acido grasso saturo composto da 16 atomi di carbonio. La necessità di sintetizzare acidi grassi emerge quando la cellula dispone di un surplus di ATP, trasformando l'acetil-CoA in macromolecole di riserva energetica. La sintesi avviene nel citoplasma, principalmente nelle cellule del fegato e del tessuto adiposo.
L'acetil-CoA, tuttavia, viene sintetizzato all'interno dei mitocondri. Pertanto, affinché la sintesi degli acidi grassi possa avvenire nel citoplasma, l'acetil-CoA deve essere trasportato fuori dal mitocondrio. Poiché l'acetil-CoA non può attraversare spontaneamente la doppia membrana mitocondriale, la cellula impiega un meccanismo astuto: l'acetil-CoA viene convertito in acido citrico, una molecola in grado di superare questa barriera attraverso un processo definito "navetta del citrato". Questa conversione è catalizzata dall'enzima citrato sintasi, che utilizza acetil-CoA e ossalacetato come substrati.
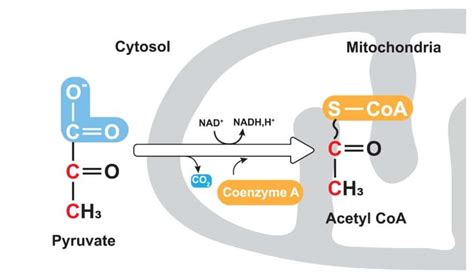
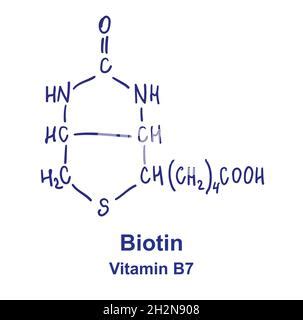
Una volta nel citoplasma, l'acetil-CoA viene sottoposto a carbossilazione, un processo che aggiunge una molecola di CO2. Questa reazione è catalizzata dall'enzima acetil-CoA carbossilasi (ACC), un passaggio chiave e altamente regolato nella biosintesi degli acidi grassi. L'ACC è un enzima ATP-dipendente e il suo funzionamento è strettamente legato alla presenza del cofattore biotina (nota anche come vitamina H o B7).
La biotina è essenziale per l'attività dell'acetil-CoA carbossilasi, legandosi covalentemente al gruppo ε-aminico di una lisina all'interno del sito attivo dell'enzima. Questo legame è cruciale per la reazione di carbossilazione, che converte l'acetil-CoA in malonil-CoA. La reazione di formazione del malonil-CoA è virtualmente irreversibile, essendo fortemente esoergonica, e consuma una molecola di ATP, liberando ADP e fosfato inorganico.
L'ossalacetato, necessario per la formazione del citrato e quindi per il trasporto dell'acetil-CoA nel citoplasma, deve essere rigenerato nel mitocondrio. Per fare ciò, viene convertito in malato nel citoplasma, utilizzando NADH e l'enzima malato deidrogenasi citosolica. Il malato può quindi rientrare nel mitocondrio, dove viene riconvertito in ossalacetato, producendo anche NADH. Un'altra via metabolica importante che coinvolge il malato nel citoplasma è quella catalizzata dall'enzima malico. Questo enzima, operando con NADP+, trasforma il malato in piruvato, producendo NADPH. Il NADPH è un cofattore riducente fondamentale, utilizzato in abbondanza durante la sintesi degli acidi grassi, che è un processo riduttivo. Pertanto, la via dell'enzima malico rappresenta una fonte significativa di NADPH per la biosintesi degli acidi grassi, oltre alla via dei pentoso fosfati.
Per ogni molecola di acetil-CoA trasportata nel citoplasma e utilizzata per la sintesi degli acidi grassi, viene prodotta una molecola di NADPH utile. Ad esempio, per sintetizzare una molecola di acido palmitico, sono necessari ben sette NADPH.
La sintesi vera e propria degli acidi grassi avviene attraverso un complesso multienzimatico chiamato acido grasso sintasi (FAS). Negli eucarioti, il FAS è un'unica grande proteina polipeptidica che contiene tutte le attività enzimatiche necessarie. Nei procarioti, invece, il sistema è composto da diversi polipeptidi. Il processo di elongazione della catena dell'acido grasso avviene attraverso reazioni cicliche.
Il primo passaggio di ogni ciclo di elongazione è la condensazione dell'acetil-ACP (derivato dall'acetil-CoA) e del malonil-ACP (derivato dal malonil-CoA) per formare acetoacetil-ACP. Successivamente, l'acetoacetil-ACP viene ridotto a D-3-idrossibutirril-ACP utilizzando NADPH. Il D-3-idrossibutirril-ACP viene poi disidratato per formare crotonil-ACP, e infine ridotto nuovamente con NADPH per produrre butirril-ACP. Queste quattro reazioni (condensazione, riduzione, disidratazione, riduzione) completano un ciclo di elongazione.
Nel ciclo successivo, il butirril-ACP formato condensa con una nuova molecola di malonil-ACP, allungando la catena di due atomi di carbonio. Questo processo si ripete ciclicamente fino alla formazione di un acido grasso a 16 atomi di carbonio (acido palmitico). Al termine di sette cicli complessivi, l'intermedio C16-acil-ACP viene scisso da una tioesterasi, liberando il palmitato e rigenerando l'ACP (Acyl Carrier Protein), una proteina fondamentale che trasporta gli intermedi della biosintesi degli acidi grassi attraverso i vari stadi enzimatici.
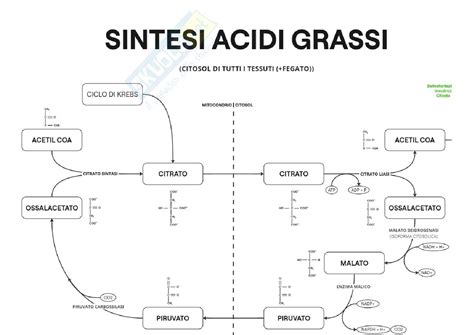
La reazione complessiva per la sintesi del palmitato è la seguente:
8 acetil-CoA + 7 ATP + 14 NADPH + 6 H+ → palmitato + 8 CoA + 7 ADP + 7 Pi + 6 H2O + 14 NADP+
Questo processo è energeticamente costoso, richiedendo sia ATP che NADPH. Il trasporto dell'acetil-CoA dal mitocondrio al citoplasma, ad esempio, consuma ATP. Inoltre, la sintesi degli acidi grassi è finemente regolata sia a livello ormonale che allosterico. L'insulina favorisce la sintesi degli acidi grassi, mentre il glucagone la inibisce. Il citrato, prodotto nel mitocondrio e trasportato nel citoplasma, agisce come attivatore allosterico dell'acetil-CoA carbossilasi, promuovendo la sintesi quando c'è abbondanza di substrati energetici. Al contrario, il palmitoil-CoA, il prodotto finale della sintesi, agisce come inibitore allosterico, segnalando alla cellula che la sintesi non è più necessaria.
La biotina, quindi, gioca un ruolo insostituibile come cofattore dell'acetil-CoA carbossilasi, l'enzima che dà inizio alla sintesi degli acidi grassi. La sua importanza si estende anche ad altre carbossilasi essenziali per il metabolismo di aminoacidi e per la gluconeogenesi, evidenziando il suo ruolo centrale nel mantenimento dell'equilibrio metabolico.
Il Ruolo della Biotina nel Metabolismo
La biotina è una vitamina idrosolubile del gruppo B, fondamentale per diverse reazioni metaboliche essenziali. Nella sua forma biologicamente attiva, è legata ai siti attivi di quattro enzimi carbossilasi:
- Acetil-CoA carbossilasi (ACC): catalizza la formazione di malonil-CoA, precursore nella sintesi degli acidi grassi.
- Piruvato carbossilasi: interviene nella gluconeogenesi, la sintesi di glucosio a partire da precursori non glucidici.
- Metilcrotonil-CoA carbossilasi: catalizza un passaggio nel metabolismo della leucina, un aminoacido essenziale.
- Propionil-CoA carbossilasi: essenziale nel metabolismo di aminoacidi, colesterolo e acidi grassi.
La biotina si trova in numerosi alimenti, con le fonti più ricche che includono il tuorlo d'uovo, il fegato e il lievito. Una parte della biotina circolante deriva anche dalla flora intestinale.
Nonostante la carenza di biotina sia rara, può manifestarsi in casi di nutrizione parenterale prolungata senza integrazione, o a seguito del consumo cronico di albumi d'uovo crudi, a causa della presenza dell'avidina, una proteina che lega la biotina impedendone l'assorbimento. I sintomi di carenza includono perdita di capelli, eruzioni cutanee e, nei casi più gravi, sintomi neurologici.
La biotina svolge un ruolo anche nella gestione del glucosio. Studi suggeriscono che pazienti con diabete mellito non-insulino-dipendente (NIDDM) presentano livelli plasmatici di biotina più bassi. La supplementazione di biotina ha dimostrato di poter ridurre i valori di glicemia a digiuno e di stimolare la sintesi di glicogeno.
Inoltre, è stato osservato che la biotina può migliorare la salute delle unghie, aumentandone lo spessore e la resistenza.
Lipogenesi sintesi acidi grassi biochimica semplificata 👌
La regolazione dell'acetil-CoA carbossilasi è complessa e coinvolge sia il rapporto insulina/glucagone che meccanismi allosterici. L'insulina promuove la defosforilazione e l'attivazione dell'ACC, favorendo la sintesi degli acidi grassi. Al contrario, il glucagone, attraverso la cascata AMPc, induce la fosforilazione dell'ACC, inattivandola e bloccando la sintesi lipidica. Il citrato agisce come attivatore allosterico dell'ACC, mentre il palmitoil-CoA agisce come inibitore.
Esistono due isoforme dell'enzima Acetil-CoA carbossilasi (ACC1 e ACC2) con localizzazioni e funzioni leggermente diverse. Si ipotizza che ACC2 sia principalmente coinvolta nella regolazione dell'ossidazione degli acidi grassi.
In sintesi, la biotina è un cofattore indispensabile per l'acetil-CoA carbossilasi, enzima chiave nella sintesi degli acidi grassi. Il suo ruolo si estende a diverse funzioni metaboliche, sottolineando la sua importanza per la salute generale dell'organismo.